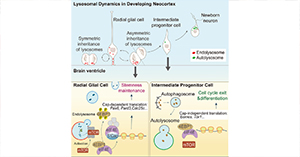
动物研究所张勇研究组及合作者揭示灵长类特异基因参与癌症及胚胎期大脑的细胞周期过程
杜布赞斯基的名言“Nothing in biology makes sense except in the light of evolution”总结了进化论对生命科学各领域的指导意义 [1]。以癌症生物学为例,进化生物学的思想与方法被大量应用[2, 3];返祖假说(atavism)及其相关研究是该领域近年兴起的一个新方向 [4, 5]。该假说认为癌症是多细胞生命逆转为其单细胞祖先的演化过程,具体表现为单细胞祖先时期即已起源基因 (unicellular gene, UC) 的表达上调和早期后生动物演化时期起源基因 (early metazoan, EM )的下调驱动癌症发生 [6, 7]。另一个探索的相对较少的假说是拮抗多效性 (antagonistic pleiotropy),进化中适应性选择近期改变的基因可使年幼或年轻的个体适合度最大化;作为代价,这些突变可能引发老年疾病,包括癌症 [8, 9]。若两种假说都成立,可以预测适应性选择所固定的人类基因组中新近产生的灵长类特异基因(primate-specific gene, PSGs)应整体表现出促进癌症发生的功能,尤其是其中衍生于UC基因的灵长类特异的重复基因更是如此。对这一联合预测的检验不仅有利于理解癌症的易感性,鉴定致癌基因;同时有助于理解PSGs如何推动适应性表型的演化。
中国科学院动物研究所张勇研究组及合作者通过整合癌症基因组图谱计划(The Cancer Genome Atlas, TCGA)数据、大脑发育转录组 [10]和灵长类特异基因集合 [11]等数据探究上述问题,相关成果于2022年12月6日在Genome Biology(GB)发表,论文标题为“Pan-cancer surveys indicate cell cycle-related roles of primate-specific genes in tumors and embryonic cerebrum”(论文链接:https://doi.org/10.1186/s13059-022-02821-9)。
为获取较为全面的基因年龄数据,研究者整合基于两种基因年龄推断策略的结果(Fig.1A):基于蛋白质家族的策略(适用于推断UC和EM基因年龄)和基于基因组共线性的策略(适用于推断PSGs基因年龄)。PSGs集合中组织特异表达基因比例明显上升;与返祖假说一致,跨癌种(pan-cancer)转录组分析显示UC和EM在癌症中分别上调和下调。PSGs同样倾向于在癌症样本中表达上升,这一结果符合拮抗多效性假说的预测(Fig.1B)。值得注意的是,PSGs上调并非由组织特异表达基因单独贡献,广谱表达的PSGs同样在癌症中表达上升(Fig.1C)。研究者通过鉴定跨癌种上调基因和跨癌种下调基因,发现PSGs富集跨癌种上调基因,UC基因表现出类似的趋势,而EM基因趋势相反(Fig.1D)。
为探究上调PSGs的功能,研究者整合了生存分析,通路富集分析和癌症细胞系基因必需性分析等多种手段。首先,生存分析显示,跨癌种上调基因uPSGs与uUC均与不利的临床预后高度关联,即高表达时倾向于促进肿瘤;而跨癌种下调基因dEM与之相反(Fig.2A)。生物通路富集分析暗示uUC和uPSGs均显著富集于细胞周期相关功能(Fig.2B);且与返祖理论相呼应,参与细胞周期的uPSGs多为UC基因的衍生拷贝(Fig.2C)。最后研究者通过关联显著影响细胞系增殖的必需基因信息,优先筛选出具备促癌功能的15个uPSGs(Fig.2D-E);其中与细胞周期功能相关的DDX11基因表现出广谱促癌功能(Fig.2F)。
DDX11参与姐妹染色单体凝聚等细胞周期过程。研究者通过进化分析推断其在灵长类演化过程中发生剧烈变化。首先,基于共线性信息,研究者发现人类基因组现在注释的DDX11为基因重复事件产生的衍生功能拷贝(Fig.3A),其旁系同源拷贝已假基因化(Fig.3B)。衍生拷贝DDX11蛋白在特定结构域呈现加速进化的情况(Fig.3C),且表达相对于外群上调(Fig.3D),暗示其功能层面的适应性变化。
根据拮抗多效性假说,促进癌症发生的细胞周期相关PSGs在适应性选择下参与生物学功能。考虑到灵长类(尤其是人类)大脑皮层扩张中伴随着神经祖细胞大量增殖,推测行使细胞周期功能的PSGs介入上述生物过程。研究者通过对大脑发育过程转录组数据的分析发现PSGs集中于最早的胚胎脑发育时期(孕后4-7周),包括DDX11在内的细胞周期相关uPSGs高表达于这一时期。与DDX11的正向选择信号相符,该胚胎发育阶段显著富集编码序列或启动子区域经历正选择的基因(Fig.4)。
该工作通过整合功能基因组和进化基因组数据发现在肿瘤中广泛上调的PSGs倾向于发挥细胞周期相关功能,且该类基因主要是UC基因的衍生重复拷贝。上述结果与返祖和拮抗多效性假说的联合预测一致:近期进化中加入遗传网络的细胞周期相关的灵长类特异基因在胚胎脑时期的高表达介导大脑皮层的扩张;作为代价,这些基因也增加了罹患癌症的风险。换言之,返祖和拮抗多效性共同贡献了癌症易感性;新基因起源、老基因调控或蛋白序列的改变共同导致了胚胎脑发育阶段细胞周期程序的系统性调整。
张勇研究员长期致力于新基因起源研究,曾揭示转座及串联重复过程如何介导新基因起源(Genome Research 2016、Nature Communications 2021、Nature Ecology & Evolution 2022);GB这项最新成果则与(Genome Research 2019)、(Developmental Cell 2021)等一起组成了该团队关于新基因如何推动人类表型演化的系列性探索。该研究由动物研究所和北京航空航天大学等机构合作完成。动物研究所博士生马辰宇、北航医学科学与工程学院副研究员李春燕及动物研究所博士后马慧静为共同第一作者,张勇研究员为通讯作者。余大奇、张宇飞、张丹、苏天晗、吴健民、王晓月、张力、陈春龙等合作者在文章写作、实验和计算分析等方面提供了大力支持。该研究得到了国家重点研发计划(2019YFA0802600)、中国科学院(XDPB17, ZDBS-LY-SM005, XBZG-ZDSYS-201913)、国家自然科学基金(31970565, 91731302)、北京脑研究中心开放研究课题等项目资助。
论文链接:https://doi.org/10.1186/s13059-022-02821-9
1. Dobzhansky T: Nothing in Biology Makes Sense except in the Light of Evolution. The American Biology Teacher 1973, 35:125-129.
2. Greaves M: Darwinian medicine: a case for cancer. Nature Reviews Cancer 2007, 7:213-221.
3. Wu C-I, Wang H-Y, Ling S, Lu X: The Ecology and Evolution of Cancer: The Ultra-Microevolutionary Process. Annual Review of Genetics 2016, 50.
4. Davies PC, Lineweaver CH: Cancer tumors as Metazoa 1.0: tapping genes of ancient ancestors. Phys Biol 2011, 8:015001.
5. Vincent MD: Cancer: beyond speciation. Adv Cancer Res 2011, 112:283-350.
6. Chen H, Lin FQ, Xing K, He XL: The reverse evolution from multicellularity to unicellularity during carcinogenesis. Nature Communications 2015, 6.
7. Trigos AS, Pearson RB, Papenfuss AT, Goode DL: Altered interactions between unicellular and multicellular genes drive hallmarks of transformation in a diverse range of solid tumors. Proceedings of the National Academy of Sciences of the United States of America 2017, 114:6406-6411.
8. Williams GC: Pleiotropy, Natural-Selection, and the Evolution of Senescence. Evolution 1957, 11:398-411.
9. Chen H, Li CY, Zhou ZC, Liang H: Fast-Evolving Human-Specific Neural Enhancers Are Associated with Aging-Related Diseases. Cell Systems 2018, 6:604-611.
10. Cardoso-Moreira M, Halbert J, Valloton D, Velten B, Chen C, Shao Y, Liechti A, Ascencao K, Rummel C, Ovchinnikova S, et al: Gene expression across mammalian organ development. Nature 2019, 571:505-509.
11. Shao Y, Chen CY, Shen H, He BZ, Yu DQ, Jiang S, Zhao SL, Gao ZQ, Zhu ZL, Chen X, et al: GenTree, an integrated resource for analyzing the evolution and function of primate-specific coding genes. Genome Research 2019, 29:682-696.

Fig.1 基因起源年龄推断及其在癌症中表达模式分析。(A) 基因起源年龄分组图。红色(9-14分枝)基于共线性策略产生,其余分枝基于蛋白质家族策略产生。(B) 14个年龄组基因在癌症与对应正常组织中表达丰度对比图。(C) 广谱表达PSGs和组织特异表达PSGs在癌症样本中表达差异统计图。(D) 跨癌种上调和下调基因的比例比较。

Fig.2跨癌种上调的PSGs(uPSGs)功能分析。(A) 与dEM基因相比,uPSGs或uUC基因的较高表达常与较短的生存时间关联。(B) 细胞周期功能相关基因在不同类型基因集合中的比例。(C) PSGs中被推断为UC 基因衍生拷贝的比例。(D) 至少被一种癌症所需的15个uPSGs的热图。(E) 对基因存在依赖的细胞系数目分布图。(F) 基因跨细胞系共同依赖性打分的密度分布图。

Fig.3 DDX11进化分析。(A). 五种灵长类动物的共线性图。(B) DDX11旁系同源基因积累功能丢失突变。(C) DDX11各功能结构域Ka/Ks统计。(D)DDX11在不同物种间表达水平比较。

Fig. 4 大脑发育不同时期偏好表达基因比例分布图。功能描述对应该大脑发育阶段高表达基因显著富集的生物学通路。
人类活动和气候变化加速生物多样性的减少,导致物种范围的转移、收缩和扩张。在全球范围内,人类活动和气候变化已对生物多样性构成了严重威胁,目前已导致全球522种灵长类动物中约68%的物种面临灭绝风险。

植物病毒素有“植物顽疾”之称,每年引起全球作物经济损失高达4000亿元。水稻作为人类重要的粮食作物之一,供给全世界一半以上的人口,主要种植于亚洲、非洲和南美洲等地区。






