白明团队利用创新形态技术合作填补羽毛演化空白
羽毛是鸟类征服天空的核心工具。羽毛演化经历了从恐龙简单的单根细丝状羽毛,逐渐变成复杂的羽片状羽毛,这个过程一般认为分为五个过渡阶段,每一个过渡阶段都是发展演化新特征或新的生长机制的结果。而羽毛的功能也从早期单根细丝状羽毛的保温、隔热、通讯交流、保护色、防水等,最终演化为飞羽的空气动力学功能。现代鸟类飞羽之所以具有高效的空气动力功能,主要在于其精密构造:首先是远端羽小枝腹侧的羽小钩嵌入隔壁羽枝上近端羽小枝背侧的凹槽,形成类似拉链的互锁结构,称为钩槽联锁机制(hook-groove mechanism);其次,近端羽小枝末端还具有结节,从而防止羽小钩在受外力时在羽小枝上滑动,称为级联滑锁系统(slide-lock system)。通过钩槽联锁机制与级联滑锁系统双重机制紧密结合,使飞行羽毛兼具超强抗撕裂能力与自修复功能。
近年来,在我国辽宁西部、河北北部和内蒙古东南部中生代地层发现的一系列带羽毛恐龙化石为解决羽毛的宏观演化提供了诸多证据。然而,由于羽毛的羽小枝等细微结构往往在成岩时被破坏,这些在页岩中保存的化石仅能保存羽毛的宏观形态。鸟类飞羽的钩槽联锁机制与级联滑锁系统如何从更原始的羽毛演化而来,始终缺乏直接证据。与此同时,中生代琥珀中保存的羽毛具有保存三维超微结构的潜力,然而如何可靠获得羽小枝微纳尺度三维超微结构,是长期存在的技术难题,这也严重阻碍了鸟类羽毛演化模型理论的进一步发展。
针对上述学科困境,中国科学院动物研究所白明团队联合国家自然博物馆、中国科学院南京地质古生物研究所、中国科学院古脊椎动物与古人类研究所、沈阳师范大学和首都师范大学等单位,对5块晚白垩世(约9900万年前)的缅甸羽毛琥珀进行研究,研究团队构建了一套针对羽毛琥珀微纳结构原位暴露和三维形态获取分析技术体系。首先,使用片溶法对羽毛化石表面进行局部处理,将羽小枝及其黑素体原位暴露。其次,利用激光共聚焦显微镜获得三维结构信息,最后,利用高分辨率扫描电镜获得羽小枝和黑素体的微纳尺度超微结构信息,更重要的是利用扫描电镜的聚焦离子束技术,对羽小枝进行原位切割,从而获得了羽小枝横截面的三维超微结构(图1)。研究结果表明,中生代羽毛的羽小枝存在两种新类型。(1)类型I羽毛,具有近乎对称的羽片,羽轴极细,羽枝基部宽度甚至小于分生的羽小枝,这一羽毛很可能属于驰龙类或伤齿龙类等接近鸟类的非鸟恐龙。然而,其羽小枝基部的横截面呈现独特的“L”形弯曲,与现代鸟类的廓羽相似,但缺乏钩槽结构。更令人惊讶的是,羽小枝中远段保留了绒羽典型的结节结构。这也证明了羽毛从绒羽向飞羽的演化是通过“基部形态改造”与“末端功能强化”的分阶段创新实现。(2)类型II羽毛,其增厚的羽轴和不对称羽片符合现代飞羽标准,这一羽毛很可能属于缅甸晚白垩世常见的反鸟类。近端羽小枝背侧首次发现明确凹槽,与远端羽小钩形成互锁机制。但与现代鸟类不同,其近端羽小枝腹侧呈“J”形弯曲,导致相邻羽小枝间存在空隙,空气易从缝隙穿过,暗示早期飞行羽毛的空气动力学效率较低。此外,研究团队还在一件无羽轴的游离羽枝标本的近端羽小枝末端背侧发现结节,这是迄今最古老的羽毛级联滑锁系统实证。
基于新发现,研究团队丰富了羽毛演化模型理论中的第3-5阶段(图2)。首先,阶段IIIa+b代表羽轴极细、羽小枝具结节但基部“L”形化的过渡形态。其次,阶段IVa显示羽轴增厚但无羽小钩;阶段IVb揭示羽轴增厚,出现羽小钩。最后,阶段Va对应反鸟类的空气动力学效率较低的不完全优化飞羽;阶段Vb则指向今鸟类的现代高效飞羽。
研究团队在观察羽毛化石超微结构的同时还发现了大量在羽小枝中呈原位保存的黑素体(图2),通过对黑素体大小及形状进行分析,同时根据黑素体的三维排列,利用布拉格定律推导的模拟公式计算其反射峰波长,首次揭示了这些羽毛可能的原始色彩:类型I羽毛因特殊黑素体层状排列可能呈现“黑色带红色光泽”;类型II羽毛为单调的深灰色或“企鹅式纯黑”;游离羽枝则为普通灰色。
该研究不仅填补了羽毛演化史的关键空白,为重建早期鸟类飞行能力的演化路径提供了珍贵实证,更启示了生物重要特征的创新往往通过“功能模块的渐进组装”实现。相关成果以“The Ultrastructure of the Cretaceous Feathers Highlights the Evolution of the Feather”为题在Science Bulletin上于近期在线发表。中国科学院动物研究所博士后王佳佳为该文第一作者。国家自然博物馆裘锐副研究员和中国科学院动物研究所白明研究员为论文共同通讯作者。该研究得到国家自然科学基金委、中国博士后管理委员会、中国科学院动物研究所、北京市、科技部和动物多样性保护与有害动物防控全国重点实验室的项目资助。
论文链接:https://doi.org/10.1016/j.scib.2025.04.073

图1 研究的类型I羽毛化石(a-e)、类型II羽毛化石(f-j)、零散羽枝化石(k-o,p-q)和现代鸟的羽毛。羽毛外形(a,f,k,p);羽小枝细节(b显示为节状羽小枝,g,l,q的白箭头指示羽小钩);羽枝外形(d,i,n);两侧羽枝截面(c,e,h,j,m,o)。比例尺:(j) 3 μm;(e)和(o) 4 μm;(c)和(h) 5 μm;(m) 10 μm;(b)和(g) 20 μm;(n) 40 μm;(k),(l),(s)-(u) 50μm;(d) 100 μm;(i),(p),(q)和(r) 200 μm;(a) 500 μm;(f) 2000 μm。
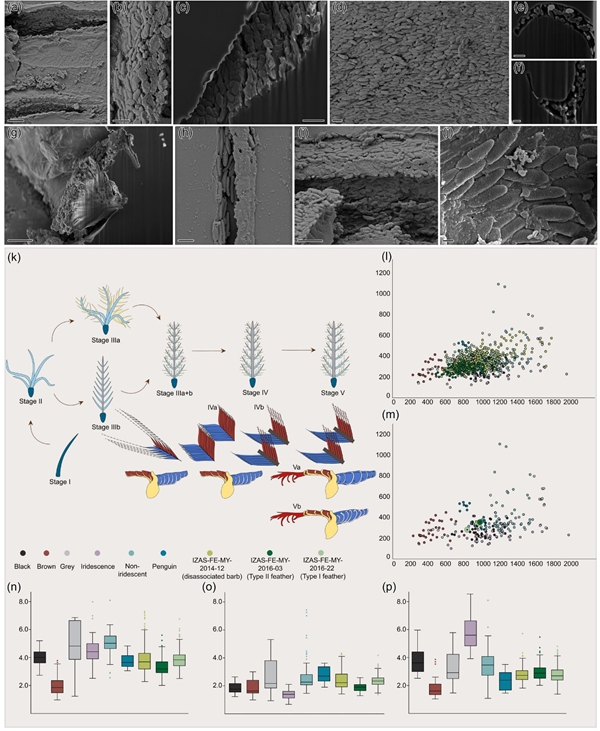
图2:羽毛化石的羽小枝中原位保存的黑素体(a-j),羽毛演化的修订模型(k)以及羽毛黑素体大小形状与其它颜色羽毛的的对比(l-p)。比例尺:(j) 0.3 μm;(f) 0.5 μm;(b),(c),(e)和(h) 1 μm;(i) 2 μm;(a)和(g) 4 μm;(d) 10 μm。
鞘翅目昆虫(甲虫)的后翅是其主要的飞行器官。研究甲虫后翅的形态特征,对于揭示其飞行机制及后翅结构的进化规律具有重要意义。然而,传统的研究方法受限于样本采集难度大、数据标注成本高等问题,阻碍了翅膀形...

大约 80% 的植物病毒依赖媒介昆虫进行传播,媒介昆虫体内的病毒稳态依赖于病毒载量与昆虫免疫系统之间的动态平衡,从而确保虫媒的生存和病毒的高效传播。

肝脏需要大量功能细胞和复杂血管网络来执行基本的生理功能,因此生物3D打印高细胞密度(约10^8细胞/mL)的肝组织在急性肝衰竭治疗中显示出巨大的应用前景。






